თავი 2. ნოციცეპცია - ტკივილის მგრძნობელობა - WEBSITE X5 UNREGISTERED VERSION 12.0.9.30 - d_training
Главное меню:
ექიმებისთვის, სტუდენტებისთვის > ნაწილი 1. ზოგადი, შესავალი
თავი 2
ნოციცეპცია - ტკივილის მგრძნობელობა
ირინა კვაჭაძე
ტკივილის სისტემა წარმოდგენილია ტკივილის ანალიზატორით. ტკივილის ანალიზატორი უზრუნველყოფს მტკივნეულ შეგრძნებათა (ტკივილის) ფორმირებას, რასაც სასიგნალო ფუნქცია აქვს დამაზიანებელი ფაქტორის მოქმედების შესახებ. მიღებულია აზრი, რომ ტკივილი შეგრძნებაა, თუმცა, ბოლო დროს მკვლევარები თვლიან, რომ ტკივილი ფსიქოფიზიოლოგიური მდგომარეობაა, რომელსაც თან ახლავს სხვადასხვა ორგანოს და სისტემის მოქმედების შეცვლა, ემოციებისა და მოტივაციების აღმოცენება. მას დიდი მნიშვნელობა აქვს დაავადებათა გამოვლინების და მკურნალობის თვალსაზრისით. ტკივილი უსიამოვნო, ტანჯვის (წვალების) სახის შეგრძნებაა (მაინც ასე ვთქვათ!), რომელიც აღმოცენდება მეტისმეტად ძლიერი გაღიზიანების, ქსოვილების დაზიანების, პათოლოგიური პროცესის განვითარების ან ქსოვილების “მჟავური შიმშილის” შედეგად.
ტკივილის შეგრძნებაზე ორგანიზმის რეაქციაში გამოჰყოფენ ნეგატიურ ემოციურ კომპონენტს, ვეგეტატურ კომპონენტს (ვლინდება შინაგანი ორგანოების მოქმედების ცვლილებებით), მოტორულ კომპონენტს (ვლინდება ტკივილის გამომწვევი პირობების ლიკვიდაციის, დაზიანებული ორგანოს დაზოგვისაკენ და სხვ. მიმართული ქცევითი აქტებით).
ტკივილის სისტემის პერიფერიული განყოფილება და ნოციცეპტორები. მოსაზრება, რომ ტკივილი “ნებისმიერი სენსორული რეცეპტორის ჭარბი გაღიზიანებისას” აღმოცენდება დღეს უარყოფილია; მიღებულია დამაჯერებელი მტკიცებულებები, რომ ტკივილი აღმოცენდება გამღიზიანებლების ზემოქმედებით სპეციალიზებულ, მაღალზღურბლოვან ტკივილის რეცეპტორებზე - ნოციცეპტორებზე (ნოცირეცეპტორებზე; ლათ. nocens – მავნე).
ჰისტოლოგიურად კანის, ლორწოვანი გარსების, ფასციების, მყესების, ჩონჩხის კუნთების, ძვლისსაზრდელას, სასახსრე ზედაპირების, წვრილი სისხლძარღვების და ლიმფური ძარღვების ადვენტიციის, ნერვული ღეროების ენდონევრიუმის, ტვინის გარსების, შინაგანი ორგანოების ქსოვილების ნოციცეპტორები ნერვული ბოჭკოების თავისუფალი, არაინკაფსულირებული დაბოლოებებია, რომლებსაც ბეწვის, წნულის, სპირალის, ან ფირფიტის ფორმა აქვთ.
ნოციცეპტორები დიდი რაოდენობითაა კანის ზედაპირზე. მათი რიცხვი ბევრად აღემატება შეხების, სითბოს და სიცივის შემგრძნები რეცეპტორების რაოდენობას. ადამიანს კანის ნოციცეპტორების უდიდესი სიმჭიდროვე აღენიშნება საზარდულის მიდამოში – 100-200 1 სმ2-ზე; ხელის გულზე, ცხვირის წვერზე, ყურის ზედაპირზე ისინი ნაკლებია – სულ 40 1 სმ2 ზე. ძალზე ბევრი თავისუფალი ნერვული დაბოლოებაა კბილის პულპაში. შემთხვევითი როდია, რომ ადამიანის კბილის პულპის არც ისე ძლიერ ელექტრულ ან მექანიკურ სტიმულაციას თან ახლავს ტკივილის შეგრძნება. მთლიანობაში, კბილის ქსოვილში ნოციცეპტორების რაოდენობა დიდია: 1 სმ2-ზე გადაანგარიშებით, დენტინისა და მინანქრის შეხების საზღვარზე დაახლოებით 75000 ნოციცეპტორია. აღსანიშნავია, პირის ღრუს ლორწოვან გარსზე არის მონაკვეთი, სადაც ნოციცეპტორები არ არის - ლოყის შიგნითა ზედაპირზე მეორე ძირითადი კბილიდან პირის კუთხემდე, ვიწრო ზოლის სახით.
ადამიანის სხვადასხვა ქსოვილოვანი სტრუქტურის ტკივილის რეცეპტორების ძირითადი ნაწილი მულტისენსორულ/პოლიმოდალური ნოციცეპტორებია:
კანის მექანოთერმონოციცეპტორები, კანქვეშა მექანოთერმონოციცეპტორები, კუნთების და შინაგანი პარენქიმული ორგანოების მექანოთერმონოციცეპტორები. იშვიათია მონოსენსორული/მონომოდალური ნოციცეპტორები: მექანონოციცეპტორები, თერმონოციცეპტორები. “სწრაფი” ტკივილი, ჩვეულებრივ, გამოიწვევა მექანიკური და თერმული მტკივნეული გამღიზიანებლებით, “ნელი” ტკივილი კი – სამი სახის მტკივნეული გამღიზიანებლით: მექანიკურით, თერმულით და ქიმიურით.
ნოციცეპტორების რეცეპტორული პოტენციალის აღმოცენების მექანიზმი საკმარისად შესწავლილი არ არის. ვარაუდობენ, რომ ნოციცეპტორის მემბრანაზე მექანიკური ზემოქმედების შემთხვევაში რეცეპტორული პოტენციალი აღმოცენდება მემბრანის ფრაგმენტის დეფორმაციის და გაჭიმვის შედეგად, რაც ააქტიურებს Na+-ის არხებს და Na+-ის დიფუზიას ნოციცეპტორის შიგნით. შესაძლებელია რეცეპტორული პოტენციალის წარმოშობის სხვა მექანიზმის არსებობაც, კერძოდ, მექანიკური ზემოქმედებით ტერმინალური სენსორული უჯრედიდან გამოყოფილი მედიატორი ურთიერთქმედებს ნოციცეპტორის მემბრანის სპეციფიკურ ქემორეცეპტორთან და განაპირობებს რეცეპტორული პოტენციალის გენერაციას.
თერმული ზემოქმედების შემთხვევაში რეცეპტორული პოტენციალი, როგორც ჩანს, ტემპერატურის გავლენით მეტაბოლური რეაქციების ინტენსიურობის ცვლილების საპასუხოდ ტერმინალური სენსორული უჯრედიდან გამოყოფილი მედიატორის მონაწილეობით ვითარდება. მედიატორი ურთიერთქმედებს ქემორეცეპტორებთან და წარმოქმნის რეცეპტორულ პოტენციალს. ამასთან, რეცეპტორული პოტენციალის განვითარების დინამიკა, რომელიც ასახავს ქემორეცეპტორებთან მედიატორის ურთიერთქმედების სიჩქარეს და წინასწარ განსაზღვრავს აღმოცენებული მოქმედების პოტენციალების სიხშირეს, როგორც ჩანს, ნოციცეპტორზე ტემპერატურული ზემოქმედებით მოდულირდება (ა. ჰაიტონი, ჯ. ჰოლი).
ანთებითი რეაქციების დროს ადამიანს ტკივილი, ჩვეულებრივ, აღეძვრება ნოციცეპტორების აქტივაციის შედეგად ქიმიური ნივთიერებებით – ალგოგენებით. ეს უკანასკნელნი, როგორც ირკვევა, ნოციცეპტორზე და სენსორული ნეირონის გამტარ მემბრანაზე ერთგვარი ქიმიური კონტეინერის სახით მოქმედებს; მასში შედის ქსოვილური ალგოგენები, რომლებიც წარმოიქმნება ქსოვილური სტრუქტურების ნგრევის შედეგად (ჰისტამინი, სეროტონინი, პროსტაგლანდინები, კალიუმის ანიონები და სხვ.), აგრეთვე, სისხლის პლაზმის ალგოგენები, რომლებიც წარმოიქმნება უჯრედშორის სივრცეში კინინების წარმოქმნის მექანიზმის კატალიზატორი სისხლის კომპონენტების გამოსვლის შედეგად; პირველ რიგში, ეს კალიდინი და ბრადიკინინია, რომლებიც მნიშვნელოვნად ზრდის ნოციცეპტორების აქტივაციას.
აღსანიშნავია, რომ ყველა ტიპის ალგოგენი მკვეთრად თრგუნავს უჯრედულ სუნთქვას. ეს კი გვაფიქრებინებს ნოციცეპტორებზე ალგოგენების გამაღიზიანებელი მოქმედების გარკვეული საერთო მექანიზმის არსებობაზე უჯრედშიდა ჟანგვის პროცესების დათრგუნვის ფონზე დაგროვილი წყალბად-იონების კონცენტრაციის ფაქტორის მონაწილეობით.
პირველადი აფერენტული ნეირონების მედიატორი გლუტამატია, მოდულატორები – სუბსტანცია P, ვიპ.
მექანიკური ტრავმის შემდგომი ანთებისას აღინიშნება სენსიტიზაცია – მგრძნობელობის მომატება ნებისმიერი გამღიზიანებლის მიმართ (დაზიანებულ უბანთან მსუბუქი შეხებაც კი ძლიერ ტკივილს იწვევს). ითვლება, რომ გამღიზიანებლის მოქმედებისას ან ქსოვილის ანთებისას ტკივილის რეცეპტორების სენსიტიზაციის ერთ-ერთი მიზეზი მეზობელი დაუზიანებელი რეცეპტორების დეპოლარიზაციაა დაზიანებული ნერვული დაბოლოებების მიერ გამოყოფილი K+-ს იონების გავლენით; კერძოდ, K+-ს მაღალი კონცენტრაცია დაუზიანებელი რეცეპტორების ირგვლივ აბრკოლებს მათგან K+-ს დიფუზიას - ვითარდება დეპოლარიზაცია. თუმცა, დაზიანებული უჯრედებიდან გამოდის არა მხოლოდ K+-ს, არამედ სხვა იონებიც (დადებითიც და უარყოფითიც). ამიტომ, უნდა ვივარაუდოთ, დაუზიანებელი რეცეპტორების დეპოლარიზაციის მიზეზი დაზიანებული უჯრედებიდან ყველა იონის გამოსვლაა, რომელთა ჯამური მუხტი უარყოფითია.
ტკივილის სისტემის გამტარი განყოფილება წარმოდგენილია B ტიპის წვრილი მიელინიზებული ბოჭკოებით და, ძირითადად, წვრილი დემიელინიზებული C ტიპის ბოჭკოებით.
ტკივილის იმპულსაციის გადაცემაში და C ბოჭკოების მონაწილეობის მტკიცებულებები მიღებულია ცდის პირების შეგრძნებათა შეფასების პროცესში კანის ნერვების ელექტრული გაღიზიანებით, ნერვული ბოჭკოების ცალკეული ჯგუფების განსხვავებული აგზნებადობის საფუძველზე. ბოჭკოებით ტკივილის იმპულსაცია მიდის ზურგის ტვინის უკანა რქებში, სადაც ზედაპირულ შრეში (განაპირა ზონა) გადაერთვება მეორე ნეირონზე. ვარაუდობენ, რომ აქ აგზნების გადაცემას უზრუნველყოფს გლუტამინის მჟავა – ცენტრალური ნერვული სისტემის ერთ-ერთი მედიატორი. შემდგომ, მეორე რიგის ნეირონების აქსონები გადადიან საპირისპირო მხარეს და ზურგის ტვინ-თალამუსის წინა ტრაქტის შედგენილობაში მიემართებიან ტვინის ღეროსკენ. ტვინის ღეროში გავლისას ზურგის ტვინ-თალამუსის წინა ტრაქტის ბოჭკოებიდან კოლატერალები მიდის ტვინის ღეროს რეტიკულური ფორმაციის ბირთვებში, შემდეგ - თალამუსში, სადაც გადაერთვება სპეციფიკური სენსორული კომპლექსის ნეირონებზე (მესამე ნეირონი), დორსალური სვეტების ბოჭკოებთან ერთად აყალიბებს მედიალურ მარყუჟს და უზრუნველყოფს აგზნების გადაცემას პროპრიოცეპტორებიდან, აგრეთვე, სახის, კანის, სხეულის და კიდურების შეხების რეცეპტორებიდან.
ტკივილის იმპულსაციის გამტარი C ბოჭკოები, ისევე როგორც ყველა მგრძნობიარე ბოჭკო, უკანა ფესვების შედგენილობაში შედის ზურგის ტვინის უკანა რქებში. ჟელატინისებრი სუბსტანციის მიდამოში ბოჭკოების მცირე ნაწილი გადაერთვება ჩართულ ნეირონებზე, ხოლო უმეტესი ნაწილი გზას განაგრძობს და გადაერთვება უკანა რქების ნეირონებზე, რომელთა აქსონები საპირისპირო მხარეს გადადის და ზურგის ტვინ-თალამუსის ლატერალური ტრაქტის შედგენილობაში ტვინის ღეროში მიემართება. სავარაუდოდ, C ბოჭკოების დაბოლოებები, რომლებიც ზურგის ტვინის ნეირონებთან კონტაქტირებს, გამოჰყოფს ორ მედიატორს - გლუტამინის მჟავას და სუბსტანცია P-ს. გლუტამინის მჟავა განსაკუთრებით სწრაფად - რამდენიმე მილიწამში ახდენს სამიზნე უჯრედზე თავისი სპეციფიკური ეფექტის რეალიზებას, ხოლო სუბსტანცია P - ძალიან ნელა: მისი ეფექტური კონცენტრაციის მიღწევას რამდენიმე წამი ან წუთიც კი სჭირდება (ა. ჰაიტონი, ჯ. ჰოლი).
ზურგის ტვინ-თალამუსის ლატერალური ტრაქტის ბოჭკოები ტვინის ღეროს დონეზე (მეორე ნეირონები) კონტაქტირებს სხვადასხვა სტრუქტურის ნეირონებთან; მათ შორის არის წყალსადენის მიმდებარე რუხი ნივთიერება, მოგრძო ტვინის, ხიდის და შუა ტვინის რეტიკულური ფორმაციის ბირთვები. აგზნება მრავალჯერ გადაირთვება ბადებრივი ფორმაციის ნეირონებზე და შემდგომ მიემართება, უპირატესად, თალამუსის არასპეციფიკურ ბირთვებში, ნაწილობრივ - თალამუსის სპეციფიკურ ბირთვებში, ჰიპოთალამუსის და სხვა ქერქქვეშა წარმონაქმნების სტრუქტურებში.
ტკივილის იმპულსაცია სახის კანიდან, პირის ღრუს ლორწოვანი გარსიდან, პერიოდონტიდან, კბილის პულპიდან A- α (“სწრაფი” ტკივილი) და C (“ნელი” ტკივილი) ბოჭკოებით სამწვერა ნერვის შედგენილობაში მიემართება მოგრძო ტვინში, სადაც მთავრდება, ძირითადად, სპინალური ტრიგემინური ტრაქტის ბირთვის ნეირონებზე; ბოჭკოების ნაწილი მიემართება მოგრძო ტვინის რეტიკულური ფორმაციის ბირთვებთან, აგრეთვე, ნაკერის ბირთვთან.
ზურგის ტვინის ტრიგემინური ტრაქტის ბირთვის ნეირონები რამდენიმე აღმავალ ტრაქტს წარმოქმნის. ყბა-სახის მიდამოს სხვადასხვა სტრუქტურის ნოციცეპტორებზე მექანიკური გამღიზიანებლების ზემოქმედებით აღმოცენებული მათი “სწრაფი” ტკივილის იმპულსაცია გატარდება ვენტრალური და დორსალური ტრიგემინურ-თალამური ტრაქტით, რომელიც თალამუსის სპეციფიკური და, ნაწილობრივ, არასპეციფიკური ბირთვების ნეირონებზე მთავრდება.
ყბა-სახის მიდამოს ორგანოებიდან “ნელი” ტკივილის იმპულსაციის გატარებაში მთავარი როლი ტრიგემინურ-რეტიკულურ-თალამურ ტრაქტს აქვს, რომელიც უზრუნველყოფს ტკივილის იმპულსაციის გენერალიზებულ გადაცემას რეტიკულური ფორმაციის ნეირონული ბადეებით თალამუსის არასპეციფიკურ ბირთვებში (ისევე, როგორც სხვა ანალიზატორებში), აგრეთვე, თავის ტვინის ლიმბურ და ჰიპოთალამუსურ სტრუქტურებში.
ამგვარად, ადამიანის ყბა-სახის ორგანოებიდან და სხეულის სხვა ნაწილებიდან ტკივილის იმპულსაციის გატარების ორგანიზებაში პრინციპული სხვაობა არ არის.
ტკივილის სისტემის ქერქული განყოფილება. რიგი ექსპერიმენტული მონაცემების საფუძველზე აშკარაა, რომ ორგანიზმის ტკივილის მგრძნობელობის წამყვან ცენტრებს ტვინის ღეროს რეტილულური ფორმაცია, ჰიპოთალამუსი და თალამუსი წარმოადგენს. დადგენილია, რომ ტვინის ქერქული უბნების იზოლირებული დაზიანება თხემის წილის ცენტრალური ღარის სიღრმეში იწვევს ტკივილის მგრძნობელობის მდგრად დაკარგვას (ნაწილობრივ ან მთლიანად) სხეულის კონტრალატერალურ ნახევარში (რ. შმიდტი). ეს მონაცემები დაემთხვა ნეიროქირურგიული ჩარევების დროს მიღებულ შედეგებს, როდესაც ცენტრალური ღარის თხემის წილის ღრმა უბნების ლოკალური ელექტრული სტიმულაცია გამოიყენებოდა. აღმოჩნდა, რომ ამგვარ სტიმულაციას ყოველთვის თან ახლდა ტკივილის მკაფიო შეგრძნება, რასაც ადრე უარყოფდნენ. ამრიგად, ნათელია, რომ ორგანიზმის მიერ ტკივილის სრულყოფილი შეგრძნება თავის ტვინის ქერქის მონაწილეობის გარეშე შეუძლებელია. ეს დასკვნა მნიშვნელოვანია დიდი ტვინის ქერქის პრიორიტეტული როლის თვალსაზრისით ბიოლოგიურად უარყოფითი ემოციური მდგომარეობების იმ მიზანმიმართული ქცევითი აქტების კონტექსტში, რომლებიც ორგანიზმს ამგვარი მდგომარეობებისაგან ათავისუფლებენ (კ. სუდაკოვი).
დღეისათვის არსებული ინფორმაციის საფუძველზე ტკივილის ანალიზატორის ქერქული პროექციული ველების ფუნქციები შემდეგია:
პირველადი სომატურ-სენსორული პროექციული ველი (C1) უზრუნველყოფს “სწრაფი” ტკივილის აღქმას, სხეულზე მისი აღმოცენების ადგილის იდენტიფიკაციას. საინტერესოა, რომ კანის ზედაპირის იმ მონაკვეთის აბსოლუტურად ზუსტი იდენტიფიკაცია, რომელზეც მტკივნეული გამღიზიანებელი მოქმედებს, მოითხოვს პროცესში კანის ტაქტილური რეცეპტორების ჩართვასაც. ნაჩვენებია, რომ კანის ტაქტილური რეცეპტორების ჩართვის გარეშე კანზე მტკივნეული უბნის იდენტიფიკაციის მაჩვენებლები საგრძნობლად უარესდება.
C1 ველს წამყვანი როლი ენიჭება მტკივნეული გაღიზიანების საპასუხოდ ორგანიზმის დამცავი მოტორული რეაქციის სასწრაფოდ ჩართვაში. შესაბამისი მოტორული რეაქციები მორფოლოგიურად და ფუნქციურად უზრუნველყოფილია C1 ველის ნეირონების პირდაპირი სინაპსური კავშირებით მოტორული ქერქის ნეირონებთან.
“ნელი” ტკივილის შეგრძნება, ჩვეულებრივ, ცუდად იდენტიფიცირდება. ცდის პირი ან კლინიკაში მყოფი პაციენტი ხშირად იტყობინება, რომ ტკივილი აღმოცენდება “ხელში”, “ფეხში”, “კეფაში” და ა.შ. “ნელი” ტკივილის ზუსტად ლოკალიზების შეუძლებლობა და/ან სირთულე იმით აიხსნება, რომ მისი გამტარი გზები, ძირითადად, თალამუსის არასპეციფიკური ბირთვების რეტიკულური ფორმაციის მულტისინაპსური ნეირონული ჯაჭვებია, რომლებიც აგზნებას გენერალიზებულად გადასცემენ ქერქის სხვადასხვა ველში, მათ შორის - ტკივილის ანალიზატორის C1 და C2 ველებში.
მეორადი სომატოსენსორული პროექციული ველი (C2) ცენტრალური ღარის და საფეთქლის წილის ზედა კიდის საზღვარზე მდებარეობს. ამ ველის ნეირონებს ორმხრივი კავშირი აქვთ თალამუსის ბირთვებთან. ეს საშუალებას აძლევს C2 ველს, შერჩევით “გაფილტროს” თალამუსში გამავალი აგზნება, პირველ რიგში, მტკივნეული წარმოშობისა. ამგვარი გამოყოფის შემდეგ C2 ველი ჩაერთვება ტვინის სისტემური მოქმედების მრავალსტადიურ პროცესებში, რომლებიც დაკავშირებულია მეხსიერებიდან “გამოთხოვილი” ქცევითი აქტის ენგრამის ამოღებასთან, მის რეალიზაციასთან ეფექტორებზე, მიღწეული სასარგებლო შედეგის შეფასებასთან, სხვ.
ცნობისთვის:
ენგრამა (ინგლ. engram მეხსიერების ფიზიოლოგიური საფუძველია და მოიცავს ნერვული ქსოვილის
ცვლილებების (მათ შორის - ბიოქიმიური) ერთობლიობას, რომელიც ადამიანის ორგანიზმზე
სინამდვილის ასახვის შედეგების შენახვას უზრუნველყოფს.
აგზნების მექანიზმის მიხედვით განარჩევენ მექანო- და ქემონოციცეპტორებს. მექანონოციცეპტორები უპირატესად განლაგებულია კანში, ფასციებში, მყესებში, სასახსრე ჩანთებში და საჭმლის მომნელებელი ტრაქტის ლორწოვანში. ისინი A-დელტა ტიპის მიელინიანი ბოჭკოების თავისუფალი ნერვული დაბოლოებანია, აგზნების გატარების სიჩქარით 4-30 მ/წმ; ისინი რეაგირებენ აგენტის მოქმედებაზე, რომელიც იწვევს რეცეპტორის მემბრანის დეფორმაციას ან დაზიანებას ქსოვილზე ზეწოლისას ან გაჭიმვისას. ამ რეცეპტორთა უმეტესობისათვის დამახასიათებელია სწრაფი ადაპტაცია. ქემონოციცეპტორები განთავსებულია კანში და ლორწოვან გარსებში, მაგრამ პრევალირებენ შინაგანი ორგანოების წვრილი არტერიების კედლებში. ისინი C ტიპის უმიელინო ბოჭკოების თავისუფალი ნერვული დაბოლოებანია, აგზნების გატარების სიჩქარე - 0,4-2 მ/წმ; ამ რეცეპტორების სპეციფიკური გამღიზიანებლები ქიმიური ნივთიერებები - ტკივილის ჰუმორული ფაქტორები ანუ ალგოგენებია, მხოლოდ ისინი, რომლებიც, "ართმევენ" რა ჟანგბადს ქსოვილებს, არღვევენ ჟანგვით პროცესებს. განარჩევენ ქსოვილურ ალგოგენებს (სეროტონინი, ჰისტამინი, აცეტილქოლინი და სხვ.), პლაზმურ ალგოგენებს (ბრადიკინინი, პროსტაგლანდინები) - ზრდიან ქემონოციცეპტორების მგრძნობელობას, და ტაქიკინინებს - გამოიყოფა ნერვული დაბოლოებებიდან მათზე დამაზიანებელი მოქმედების დროს.
სადღეისოდ ითვლება, რომ ტკივილი შეიძლება აღმოცენდეს, ასევე, სხვადასხვა მოდალობის რეცეპტორებზე ინტენსიური ზემოქმედების დროს. ამიტომ ტერმინში "ნოციცეპტორი" იგულისხმება არა მარტო რეცეპტორული აპარატი, არამედ მასთან დაკავშირებული აფერენტული ბოჭკოს პერიფერიული ნაწილი. ამ წარმონაქმნს ეწოდა "სენსორული ტკივილითი ერთეული". ითვლება, რომ მას აქვს ორი აგზნებადი უბანი: ერთი - დენდრიტის პრეტერმინალური ნაწილი – აიგზნება მხოლოდ დამაზიანებელი სტიმულებით, მეორე – საკუთრივ ტერმინალი - შეიძლება აქტივირდეს არანოციცეპტური ინფორმაციითაც.
ტკივილის რეცეპტორების ადაპტაცია, როგორც აღინიშნა, შესაძლებელია, მაგრამ უმრავლეს შემთხვევაში ისინი მნიშვნელოვანი ადაპტაციის უნარს არ ავლენენ, რაც ანალგეზიის აუცილებლობას წარმოშობს.
ორგანიზმის ანტინოციცეპტური სისტემა (გაუტკივარების, ტკივილსაწინააღმდეგო) ტკივილის სისტემის ნაწილია.
მე-20 საუკუნის 70-იან წლებში ჩამოყალიბდა შეხედულებანი ორგანიზმში ენდოგენური ანტინოციცეპტური სისტემის შესახებ. მისი ფუნქცია ნოციცეპტური სისტემების კონტროლი და მათი ზეაგზნების პროფილაქტიკაა.
როდესაც ანტინოციცეპტური სისტემა ამ შეზღუდვის ფუნქციას ვერ ასრულებს, ვითარდება "მტკივნეული, ტკივილითი შოკი". ანტინოციცეპტური სისტემა ცნს-ის სხვადასხვა დონეზე განლაგებულ სტრუქტურათა ერთობლიობაა საკუთარი ნეიროქიმიური მექანიზმებით.
ამ ერთობლიობის პირველი დონე წარმოდგენილია შუა, მოგრძო და ზურგის ტვინის კომპლექსით (ნაკერის ბირთვი, რეტიკულური ფორმაცია, ჟელატინური სუბსტანცია). ამ სტრუქტურების აგზნება ეფერენტულად იწვევს ზურგის ტვინის "ტკივილის ჭიშკრის" შეკავებას და აღმავალი ნოციცეპტური ნაკადის დათრგუნვას. ამ დონის მედიატორებია სეროტონინი და ოპიოიდები.
მეორე დონე წარმოდგენილია, ძირითადად, ჰიპოთალამუსით: აკავებს ზურგის ტვინის ნოციცეპტურ ნეირონებს, ააქტივებს პირველი დონის ანტინოციცეპტურ მექანიზმებს, თრგუნავს თალამუსის ნოციცეპტურ ნეირონებს. ჰიპოთალამუსი "მუშაობს" ადრენერგული და ოპიოიდური ნეიროქიმიური მექანიზმებით.
მესამე დონე თავის ტვინის ქერქია, უფრო ზუსტად კი - II სომატოსენსორული ზონა. აქ ფორმირდება ანტინოციცეპტური სისტემის სხვა სტრუქტურების აქტივობა და ადეკვატური რეაქციები დამაზიანებელ ზემოქმედებაზე.
ენდოგენური ანტინოციცეპტური სისტემის შესწავლის შედეგად აღწერილია ოპიატური რეცეპტორები, რომლებიც რეაგირებენ მორფინზე და სხვა ოპიოიდებზე. ისინი აღმოჩენილია ბევრ ქსოვილში, მაგრამ ძირითადად - ცნს-ში, აფერენტული იმპულსაციის გადართვის თითქმის ყველა დონეზე. აღწერილია ოპიატური რეცეპტორების ოთხი ტიპი: მიუ-, დელტა-, კაპა- და სიგმა-რეცეპტორები.
ორგანიზმში გამომუშავდება საკუთარი ენდოგენური ოპიოიდური ნივთიერებანი - ოლიგოპეპტიდები, წოდებული ენდორფინებად (ენდომორფინებად), ენკეფალინებად და დინორფინებად. ეს ნივთიერებანი, უკავშირდება რა ოპიატურ რეცეპტორებს, იწვევს პრე- და პოსტსინაპსურ შეკავებას ნოციცეპტურ სისტემაში, რასაც შედეგად მოჰყვება ანალგეზია და ჰიპოალგეზია. ოპიატური რეცეპტორების ასეთი ჰეტეროგენულობა და ოპიოიდური პეპტიდების შერჩევითი მგრძნობელობა (აფინიტეტი) მათ მიმართ ასახავს სხვადასხვა წარმოშობის ტკივილის განსხვავებულ მექანიზმს.
ტკივილის მგრძნობელობის რეგულირებაში მონაწილეობენ არაოპიოდური ნივთიერებანიც - ნეიროტენზინი, ანგიოტენზინ II, კალციტონინი, ბომბეზინი, ქოლეცისტოკინინი, რომელნიც შემაკავებლად მოქმედებენ ნოციცეპტური იმპულსაციის გატარებაზე. ეს ნივთიერებები გამოიყოფა ცნს-ის სხვადასხვა მიდამოში და აქვთ თავისი რეცეპტორები ნოციცეპტური იმპულსაციის "გადართვის" სხვადასხვა "სადგურში". მათი ანალგეზიური ეფექტი ტკივილის ბუნებაზეა დამოკიდებული. კერძოდ, ნეიროტენზინი ბლოკავს ვისცერულ ტკივილს, ქოლეცისტოკინინი კი - თერმული გამღიზიანებლით გამოწვეულს.
ალგეზიმეტრიის (დამოკიდებულების დადგენა ორგანიზმზე მოქმედ მტკივნეულ გამღიზიანებელსა და საპასუხოდ აღმოცენებულ სუბიექტურ რეაქციას შორის) მიმდინარეობისას ნაჩვენებია, რომ ტკივილის შეგრძნება პირველად ჩნდება გამღიზიანებლის გარკვეული მინიმალური ძალის პირობებში, რომელსაც ტკივილის ზღურბლი ეწოდა. გამღიზიანებლის ძალას, რომლის შემთხვევაში ცდის პირი აუტანელი ტკივილის გამო მისი მოქმედების შეწყვეტას ითხოვს, ტკივილისადმი მდგრადობის ზღურბლი ეწოდა.
ალგეზიმეტრიით, ფსიქოლოგიური, კლინიკური დაკვირვებებით სხვადასხვა გამოკვლევის დროს ცდის პირების მიერ მტკივნეული ზემოქმედების ატანის უნარის საკითხის დეტალურმა შესწავლამ აჩვენა, რომ ცდის პირების ზოგადი მდგომარეობის შეცვლისას, ასევე, იცვლება ტკივილის ზღურბლი და ტკივილისადმი მდგრადობა. რიგ შემთხვევებში ეს ჰიპერალგეზიაა (ტკივილისადმი მგრძნობელობის მომატება), მაგრამ უფრო ხშირად - ჰიპოალგეზია (ტკივილისადმი მგრძნობელობის შემცირება) ანალგეზიამდე, ანუ ტკივილისადმი სრულ შეუგრძნებლობამდეც კი.
ცხოველებზე და ადამიანებზე ჩატარებული გამოკვლევების შედეგებმა დაადასტურა, რომ შუა და მოგრძო ტვინის, ქერქქვეშა ბირთვების სტიმულაცია იწვევს სარწმუნო ჰიპოალგეზიას ან ანალგეზიას.
კლინიკურ პრაქტიკაში გამოყენებული გაუტკივარების ფიზიოლოგიური მექანიზმები. ადგილობრივი ანესთეზია ხორციელდება მტკივნეული იმპულსაციის გატარების ბლოკადის გზით პერიფერიული ნერვული ბოჭკოების ნოციცეპტორების დონეზე. მისი ერთ-ერთი ვარიანტი ადგილობრივი ინფილტრაციული ანესთეზიაა, როდესაც ქსოვილი (მაგალითად, კუნთი) ინფილტრირდება საანესთეზიო ნივთიერებით.
ადგილობრივი ანესთეზიის ხანგრძლივობა დამოკიდებულია ანესთეტიკზე, გრძელდება 20-60 წუთს, გამოიყენება ხანმოკლე, მცირე მოცულობის ჩარევისას, მაგალითად, კბილის ექსტრაქციისას და სხვ. ადგილობრივი ანესთეტიკები (მაგალითად, ნოვოკაინი) ბლოკავს ნერვული ბოჭკოების ნატრიუმის პოტენციალდამოკიდებული არხების “ჭიშკროვან” რეაქციებს; შედეგად მოქმედების გავრცელებად პოტენციალებს ანესთეტიკით დამუშავებული ბოჭკოს ზონაში ნატრიუმის არხების აქტივაციის და რეგენერაციული დეპოლარიზაციის განვითარების უნარი აღარ შესწევთ. სწორედ ამიტომ, დამუშავებულ უბანში ნერვული ბოჭკოები დროებით კარგავენ აგზნებადობას და გამტარებლობას.
ზოგადი ანესთეზია გამოიყენება რთული, ხანგრძლივი ოპერაციული ჩარევებისას: ინჰალაციურად ან ინტრავენურად შეყვანილი სანარკოზო პრეპარატები ტკივილის იმპულსაციის გატარებას წყვეტენ, უპირატესად, ქერქქვეშა დონეზე.
ბოლო წლებში ფართო გავრცელება ჰპოვა გაუტკივარების არატრადიციულმა მეთოდებმა. მათ შორის აღსანიშნავია აკუპუნქტურა (ნემსით ჩხვლეტა) - სხეულის ბიოლოგიურად აქტიურ წერტილებზე (სენსორული რეცეპტორების თავმოყრის ადგილებზე) საგანგებო ნემსებით მექანიკური ზემოქმედების მეთოდი. აკუპუნქტურის სახესხვაობას წარმოადგენს ელექტროაკუპუნქტურა - ბიოლოგიურად აქტიურ წერტილებზე ელექტრული ზემოქმედების მეთოდი. ორივე შემთხვევაში ბიოლოგიურად აქტიური წერტილების 20-30 წუთის განმავლობაში სტიმულაცია იწვევს ანალგეზიის ეფექტს. დადგენილია, რომ აკუპუნქტურული ანალგეზიისას ადგილი აქვს ჰიპოფიზიდან სისხლში ენდორფინების რეფლექსურად გამოსროლას, შემდგომ ენდორფინები უკავშირდება ოპიატურ რეცეპტორებს და არღვევს ტკივილის იმპულსაციის სინაპსურ გადაცემას ცნს-ში (ბ. პომერანი). ამ ჰიპოთეზას ადასტურებს ვირთაგვებისათვის ჰიპოფიზის ამოკვეთით ჩატარებული ცდები: ჰიპოფიზექტომიის შემდეგ აკუპუნქტურული ანალგეზია არ ვითარდება.
ნოციცეპტური და ანტინოციცეპტური სისტემები შედიან ერთ ფუნქციურ სისტემაში, რომელიც მიმართულია ორგანიზმის ქსოვილების მთლიანობის შენარჩუნებაზე. მათი ნორმალური მუშაობა შესაძლებელია მხოლოდ ორივეს აქტივობის პირობებში. ანტი- და ნოციცეპტური სისტემების ტონუსის ცვლილებები ტკივილის მგრძნობელობის შეცვლას იწვევს. მაგალითად, ნოციცეპტური სისტემის ტონუსის მომატება, ან ანტინოციცეპტურის ტონუსის შემცირება დაძაბვის (ყურადღების დაძაბვის) რეაქციის დროს ამცირებს ტკივილის ზღურბლს (ე.ი. ზრდის მგრძნობელობას), რაც იძლევა ორგანიზმისათვის საშიში გარემო გამღიზიანებლების იდენტიფიკაციის მეტ საშუალებას.
ტკივილის შეგრძნების აღმოცენებაში დიდი მნიშვნელობა ენიჭება ჰისტამინის გამოთავისუფლებას, რომელიც კანში გამოიყოფა მისი გაღიზიანებისას, ანთების, დამწვრობის, დაჟეჟილობის და სხვა დროს. კანის დაზიანებისას მასში გამოიყოფა, აგრეთვე, ბრადიკინინი, რომლის ინტრავენური შეყვანა იწვევს ძლიერ ტკივილს. როგორც აღინიშნა, ეს ნივთიერებები ტკივილის ჰუმორულ ფაქტორები - ალგოგენებია.
ხარისხობრივი მახასიათებლების მიხედვით, ვისცერული ტკივილი ბევრი ნიშნით ემთხვევა სომატურ ტკივილს. მისი აღმოცენების ძირითადი მიზეზებია:
- ქსოვილების იშემია, რასაც მეტაბოლიტების დაგროვება და ქსოვილური სუნთქვის დარღვევა მოსდევს;
- ღრუ ორგანოების კედლების მთლიანობის დაზიანებისას მათი შიგთავსის გარეთ გამოსვლა, მაგალითად, აპენდიქსის ანთების და გახეთქვისას მისი შიგთავსის გამოსვლა მუცლის ღრუში;
- კუჭის, ნაღვლის ბუშტის, ნაწლავების, შარდის ბუშტის, შარდსაწვეთების კუნთების ძლიერი ტონური შეკუმშვა ან გაჭიმვა.
ხანგრძლივობის მიხედვით განარჩევენ მწვავე, ქვემწვავე და ქრონიკულ ტკივილს. მწვავე უცბად აღმოცენებული ტკივილია. მწვავე ტკივილს, ჩვეულებრივ, გამაფრთხილებელი, ანუ სასიგნალო მნიშვნელობა აქვს. ქრონიკული ტკივილი ხანგრძლივი, დიდხანს შენარჩუნებადი ან სისტემატურად განმეორებადი, სხვადასხვა ინტენსიურობის ტკივილია. ამგვარი ტკივილის მაგალითებია გარკვეული სახის თავის ტკივილი, ზურგის ტკივილი ოსტეოქონდროზის დროს, ტკივილი სიმსივნეების განვითარების გვიან სტადიებზე. ქვემწვავე ტკივილს, მახასიათებლების მიხედვით, ერთგვარი შუალედური პოზიცია აქვს მწვავე და ქრონიკული ტიპის ტკივილებს შორის.
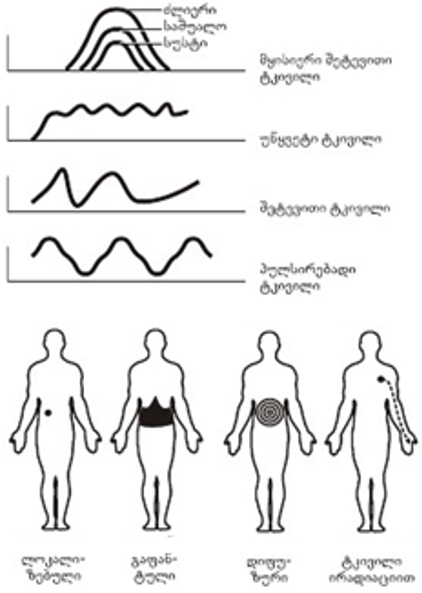
აქვე უნდა აღინიშნოს, ასევე, მყისიერი, შეტევის მაგვარი, უწყვეტი, მოპულსირე ტკივილები, რომლებიც ასახავს ტკივილის ინტენსიურობის სუბიექტურ, ხარისხობრივ აღქმას დროის გარკვეული ინტერვალის ფარგლებში.
აღმოცენების მიზეზთან მიმართებით არჩევენ ფიზიკურ და ფსიქოგენურ ტკივილს. ფიზიკური ტკივილი წარმოიშობა სხვადასხვა გამღიზიანებლის ზემოქმედებით ორგანიზმის ქსოვილთა სტრუქტურებზე. ფსიქოგენური ტკივილი ადამიანის ფსიქიკური ფუნქციის დარღვევის შედეგია.
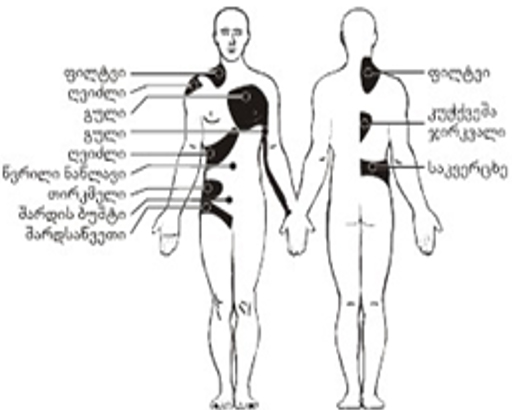
გამოჰყოფენ, აგრეთვე, არეკვლილ/ირადირებულ ტკივილს – ტკივილის შეგრძნებას ორგანიზმში მისი გამოწვევის ადგილიდან დაშორებულ ნაწილში, მაგალითად, სტენოკარდიის შეტევისას შეიძლება ტკივილი მარცხენა ხელში აღიძრას და სხვ.
ტკივილი მნიშვნელოვანი ხარისხით სუბიექტური შეფასების საგანია და სხვადასხვა "ხასიათი" შეიძლება ჰქონდეს (სურ. 1). შინაგანი ორგანოების ტკივილისას მისი ლოკალიზაცია ხშირად არამკაფიოდაა გამოხატული. შინაგანი ორგანოების დაავადებებისას ტკივილი შეიძლება შეიგრძნობოდეს არა დაავადების ადგილზე, არამედ სხეულის სხვა უბნებში, მაგალითად, კანის ზედაპირზე. ასე მაგალითად, სტენოკარდიის დროს ტკივილი აღმოცენდება არა მარტო გულის მიდამოში, არამედ, შესაძლებელია, მარცხენა ხელსა და ბეჭში, კისრის და თავის მარცხენა ნახევარში (სურ. 2).
კანის რეცეპტორების გაღიზიანების დროს აღმოცენებულ თავისებურ არასასიამოვნო შეგრძნებას წარმოადგენს ქავილი, რომელიც იწვევს კანის ფხანის რეფლექსურ რეაქციას. ქავილის შეგრძნებას უკავშირებენ ტკივილის რეცეპტორებს, რომლებიც ეპიდერმისის ქვეშ არიან განლაგებული და წარმოადგენენ თავისუფალ ნერვულ დაბოლოებებს. ქავილის წარმოშობაში მნიშვნელობა აქვს კანში ზოგიერთი ქიმიური ნივთიერების წარმოქმნას, რომლებიც აღიზიანებენ რეცეპტორებს (ჰისტამინი, პეპტიდაზები).
ილუსტრაცია